ShopDreamUp AI ArtDreamUp

Deviation Actions








Suggested Deviants









Suggested Collections
















You Might Like…












Featured in Groups





Description
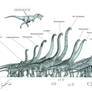
Lapparentosaurus madagascariensis
Etymology: "Lapparent's lizard from Madagascar"
Time horizon: Middle Jurassic, Bathonian epoch (~167 mya)
Length: ~24m (~79 ft.), based on adult caudal remains and scaling up proportions from immature specimens. *Updated with correct scaling for Lydekker's giant vertebrae*
Probable mass: 30+ tons
The earliest "normal" brachiosaur that we have at least semi-decent material for, Lapparentosaurus madagascariensis. Of course, exactly what is "normal" for a brachiosaur is still very much open to debate, but at least it's the most basal one that has the long thumb metacarpals typical of later species in the family. This animal is surprisingly well-represented in the fossil record, yes considering that, it's one of the most poorly researched brachiosaurs in existence. The first remains were found in the late 1800s, as British and French paleontologists rushed worldwide to discover new dinosaurs, often fragments of sauropods that were inevitably thrown without much thought into Titanosaurus, Cetiosaurus, or Bothriospondylus.
Such was the case with the material which would later become known as Lapparentosaurus. The history of "Bothriospondylus" is a long and insanely confusing one. But in a nutshell it was one of the original Dinosauria described by Owen in the heady times of the Victorian age, and the type species appears to have been a brachiosaur, or at least a basal macronarian of some sort. Other remains from England and Portugal were attributed as various species of Bothriospondylus, and over the years, due to various rivalries between Owen, Seeley, Mantell, Lydekker, Von Huene, Nopcsa and many others, the panoply of ascribed remains filtered through countless reclassifications as new forms of Cetiosaurus, Oplosaurus, Ornithopsis, Pelorosaurus, and Bothriospondylus. Most of the bones that made up this confusing tangle had no relation to each other, and could be said to represent entire Jurassic and Early Cretaceous faunas jumbled together. Some were unmistakable brachiosaurids, others were basal "cetiosaurid"-grade eusauropods, some were rebbachisaurs referred from Argentina (e.g. Nopcsaspondylus), and yet others were random bits of Iguanodon thrown in by Sir Owen for good measure and the sake of obligatory Crystal Palace imperial nationalism. Various remains, whether in England, Madagascar, or South America, were hastily referred to this genus by later paleontologists, all before modern taxonomy and cladistics had come into being. Bothriospondylus madagascariensis formed a chimera of nearly every large non-titanosaur sauropod found in Madagascar (the titanosaur remains were, not surprisingly, all thrown into Titanosaurus without much analysis either).
In Madagascar, Lydekker and his French counterpart Armand Thevenin collected most of the bones from the larger B. madagascariensis specimens currently included in Lapparentosaurus. Many of these are centra, missing most of the neural arch, as well as limb bones from a few large specimens. They do not appear to be juveniles due to remaining small parts of the neural arch being fused to the centrum. In 1943, Albert-Felix de Lapparent, a French paleontologist and Catholic priest who would later gain fame for dinosaur discoveries all over Africa and southern Europe, unearthed a far more complete specimen in France and also referred it to Bothriospondylus. This animal is a distinct brachiosaur species, and not related to the Madagascar material. In 1955, Lavocat referred a specimen which is now known to be very different from Lapparentosaurus, with no less than five carpal bones and a much more primitive thumb unit that had the short metacarpal and huge swiveling scythe-claw common in more basal macronarians, in addition to having well-developed secondary phalanges, another primitive trait. More discoveries by Collignon (1953) and de Ricqles (1968) filled in the gaps, though with material which may actually belong to yet another genus. In 1975, Annie Ogier, a PhD student in the University of Paris, both increased and frustrated our understanding of this animal when she described a plethora of juvenile remains and assigned MAA 91-92, a pair of posterior dorsal neural arches, as the holotype of B. madagascariensis (despite the fact that Lydekker had already assigned the fragmentary mid-dorsal vertebra BMNH R2598 as the holotype in 1895). Ogier's findings were only documented in her PhD thesis, which was never published and exists in only a few copies in Paris libraries. She seems to have since disappeared from the paleontological profession since then, and never published further papers, thus depriving many researchers of the photographic evidence for most of these specimens.
In 1986 José Bonaparte analyzed the B. madagascariensis specimens and redescribed most of the material, including Ogier's juvenile bones, as Lapparentosaurus, in honor of Albert-Felix de Lapparent, although Lapparent had never handled the Madagascar material. To add a further irony, he chose Ogier's MAA 91-92 as the holotype instead of Lydekker's original material (which though fragmentary, is nevertheless good enough to be diagnostic of a basal Europasaurus-grade brachiosaur).
After Lavocat's material and the French and South American specimens were thrown out in recent years, what remains is a more or less unified Lapparentosaurus at the basal end of Brachiosauridae, closest to Europasaurus, and more derived than Atlasaurus, Volkheimeria, the Klamelisaurids, and whatever Lavocat's bizarre 5-carpal bone species turns out to be. The material currently placed in Lapparentosaurus represents three main individuals and as many as seven less complete ones from multiple growth stages. What is shown here is only the material which is known from available photographs, published or otherwise, or could otherwise be reasonably figured from the specimens still referred to this taxon. This may be only a small sample of the valid Lapparentosaurus material lying in museum vaults in London, Paris, or elsewhere. Lapparentosaurus is also one of the first dinosaurs to be subjected to histological age analysis, and in 1983 it was determined that one mature individual was 45 years old, and still had not stopped growing. Sexual maturity in big sauropods thus took place long before "full size range" was reached. It was also shown in 1995 to have been relatively fast-growing given the presence of a large amount of fibrolamellar and harversian bone, which firmly established Bakker's theory that sauropods were warm-blooded.
References:
R. Lydekker, 1895, "On bones of a sauropodous dinosaur from Madagascar", Quarterly Journal of the Geological Society of London 51: 329-336
J.F. Bonaparte, 1986, "Les dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétosauridés) du Jurassique Moyen de Cerro Cóndor (Chubut, Argentina)", Annales de Paléontologie (Vert.-Invert.) 72(3): 325-386
de Lapparent, A.F., 1943, "Les dinosauriens jurassiques de Damparis (Jura)", Mémoires de la Société Géologique de France 47: 5–20
Mannion, P.D., 2010 "A revision of the sauropod dinosaur genus 'Bothriospondylus' with a redescription of the type material of the middle Jurassic form 'B. madagascariensis'", Palaeontology, 53(2): 277–296
Ogier, A., 1975. Etude de nouveaux ossements de Bothriospondylus (Sauropode) d’un gisement du Bathonien de Madagascar. Unpublished PhD Thesis, Universite´ de Paris, 102 pp.
de Ricqlès, A., 1983, "Cyclical growth in the long bones of a sauropod dinosaur", Acta Palaeontologia Polonica 28: 225-232
Rimblot-Baly, F., A. de Ricqlès, & L. Zylberberg, 1995, "Analyse paléohistologique d'une série de croissance partielle chez Lapparentosaurus madagascariensis (Jurassique Moyen): essai sur la dynamique de croissance d'un dinosaure sauropode", Annales de Paléontologie 81: 49–86
Thevenin, A. 1907. Paleontologie de Madagascar. Annales de Paleontologie, 2, 121–136.
Etymology: "Lapparent's lizard from Madagascar"
Time horizon: Middle Jurassic, Bathonian epoch (~167 mya)
Length: ~24m (~79 ft.), based on adult caudal remains and scaling up proportions from immature specimens. *Updated with correct scaling for Lydekker's giant vertebrae*
Probable mass: 30+ tons
The earliest "normal" brachiosaur that we have at least semi-decent material for, Lapparentosaurus madagascariensis. Of course, exactly what is "normal" for a brachiosaur is still very much open to debate, but at least it's the most basal one that has the long thumb metacarpals typical of later species in the family. This animal is surprisingly well-represented in the fossil record, yes considering that, it's one of the most poorly researched brachiosaurs in existence. The first remains were found in the late 1800s, as British and French paleontologists rushed worldwide to discover new dinosaurs, often fragments of sauropods that were inevitably thrown without much thought into Titanosaurus, Cetiosaurus, or Bothriospondylus.
Such was the case with the material which would later become known as Lapparentosaurus. The history of "Bothriospondylus" is a long and insanely confusing one. But in a nutshell it was one of the original Dinosauria described by Owen in the heady times of the Victorian age, and the type species appears to have been a brachiosaur, or at least a basal macronarian of some sort. Other remains from England and Portugal were attributed as various species of Bothriospondylus, and over the years, due to various rivalries between Owen, Seeley, Mantell, Lydekker, Von Huene, Nopcsa and many others, the panoply of ascribed remains filtered through countless reclassifications as new forms of Cetiosaurus, Oplosaurus, Ornithopsis, Pelorosaurus, and Bothriospondylus. Most of the bones that made up this confusing tangle had no relation to each other, and could be said to represent entire Jurassic and Early Cretaceous faunas jumbled together. Some were unmistakable brachiosaurids, others were basal "cetiosaurid"-grade eusauropods, some were rebbachisaurs referred from Argentina (e.g. Nopcsaspondylus), and yet others were random bits of Iguanodon thrown in by Sir Owen for good measure and the sake of obligatory Crystal Palace imperial nationalism. Various remains, whether in England, Madagascar, or South America, were hastily referred to this genus by later paleontologists, all before modern taxonomy and cladistics had come into being. Bothriospondylus madagascariensis formed a chimera of nearly every large non-titanosaur sauropod found in Madagascar (the titanosaur remains were, not surprisingly, all thrown into Titanosaurus without much analysis either).
In Madagascar, Lydekker and his French counterpart Armand Thevenin collected most of the bones from the larger B. madagascariensis specimens currently included in Lapparentosaurus. Many of these are centra, missing most of the neural arch, as well as limb bones from a few large specimens. They do not appear to be juveniles due to remaining small parts of the neural arch being fused to the centrum. In 1943, Albert-Felix de Lapparent, a French paleontologist and Catholic priest who would later gain fame for dinosaur discoveries all over Africa and southern Europe, unearthed a far more complete specimen in France and also referred it to Bothriospondylus. This animal is a distinct brachiosaur species, and not related to the Madagascar material. In 1955, Lavocat referred a specimen which is now known to be very different from Lapparentosaurus, with no less than five carpal bones and a much more primitive thumb unit that had the short metacarpal and huge swiveling scythe-claw common in more basal macronarians, in addition to having well-developed secondary phalanges, another primitive trait. More discoveries by Collignon (1953) and de Ricqles (1968) filled in the gaps, though with material which may actually belong to yet another genus. In 1975, Annie Ogier, a PhD student in the University of Paris, both increased and frustrated our understanding of this animal when she described a plethora of juvenile remains and assigned MAA 91-92, a pair of posterior dorsal neural arches, as the holotype of B. madagascariensis (despite the fact that Lydekker had already assigned the fragmentary mid-dorsal vertebra BMNH R2598 as the holotype in 1895). Ogier's findings were only documented in her PhD thesis, which was never published and exists in only a few copies in Paris libraries. She seems to have since disappeared from the paleontological profession since then, and never published further papers, thus depriving many researchers of the photographic evidence for most of these specimens.
In 1986 José Bonaparte analyzed the B. madagascariensis specimens and redescribed most of the material, including Ogier's juvenile bones, as Lapparentosaurus, in honor of Albert-Felix de Lapparent, although Lapparent had never handled the Madagascar material. To add a further irony, he chose Ogier's MAA 91-92 as the holotype instead of Lydekker's original material (which though fragmentary, is nevertheless good enough to be diagnostic of a basal Europasaurus-grade brachiosaur).
After Lavocat's material and the French and South American specimens were thrown out in recent years, what remains is a more or less unified Lapparentosaurus at the basal end of Brachiosauridae, closest to Europasaurus, and more derived than Atlasaurus, Volkheimeria, the Klamelisaurids, and whatever Lavocat's bizarre 5-carpal bone species turns out to be. The material currently placed in Lapparentosaurus represents three main individuals and as many as seven less complete ones from multiple growth stages. What is shown here is only the material which is known from available photographs, published or otherwise, or could otherwise be reasonably figured from the specimens still referred to this taxon. This may be only a small sample of the valid Lapparentosaurus material lying in museum vaults in London, Paris, or elsewhere. Lapparentosaurus is also one of the first dinosaurs to be subjected to histological age analysis, and in 1983 it was determined that one mature individual was 45 years old, and still had not stopped growing. Sexual maturity in big sauropods thus took place long before "full size range" was reached. It was also shown in 1995 to have been relatively fast-growing given the presence of a large amount of fibrolamellar and harversian bone, which firmly established Bakker's theory that sauropods were warm-blooded.
References:
R. Lydekker, 1895, "On bones of a sauropodous dinosaur from Madagascar", Quarterly Journal of the Geological Society of London 51: 329-336
J.F. Bonaparte, 1986, "Les dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétosauridés) du Jurassique Moyen de Cerro Cóndor (Chubut, Argentina)", Annales de Paléontologie (Vert.-Invert.) 72(3): 325-386
de Lapparent, A.F., 1943, "Les dinosauriens jurassiques de Damparis (Jura)", Mémoires de la Société Géologique de France 47: 5–20
Mannion, P.D., 2010 "A revision of the sauropod dinosaur genus 'Bothriospondylus' with a redescription of the type material of the middle Jurassic form 'B. madagascariensis'", Palaeontology, 53(2): 277–296
Ogier, A., 1975. Etude de nouveaux ossements de Bothriospondylus (Sauropode) d’un gisement du Bathonien de Madagascar. Unpublished PhD Thesis, Universite´ de Paris, 102 pp.
de Ricqlès, A., 1983, "Cyclical growth in the long bones of a sauropod dinosaur", Acta Palaeontologia Polonica 28: 225-232
Rimblot-Baly, F., A. de Ricqlès, & L. Zylberberg, 1995, "Analyse paléohistologique d'une série de croissance partielle chez Lapparentosaurus madagascariensis (Jurassique Moyen): essai sur la dynamique de croissance d'un dinosaure sauropode", Annales de Paléontologie 81: 49–86
Thevenin, A. 1907. Paleontologie de Madagascar. Annales de Paleontologie, 2, 121–136.
Image size
4240x6528px 3.34 MB
© 2013 - 2024 Paleo-King
Comments47
Join the community to add your comment. Already a deviant? Log In
So was Madagascar an island at the time. That leaves interesting implications for Island fauna.